地球氧化大气层的未来寿命
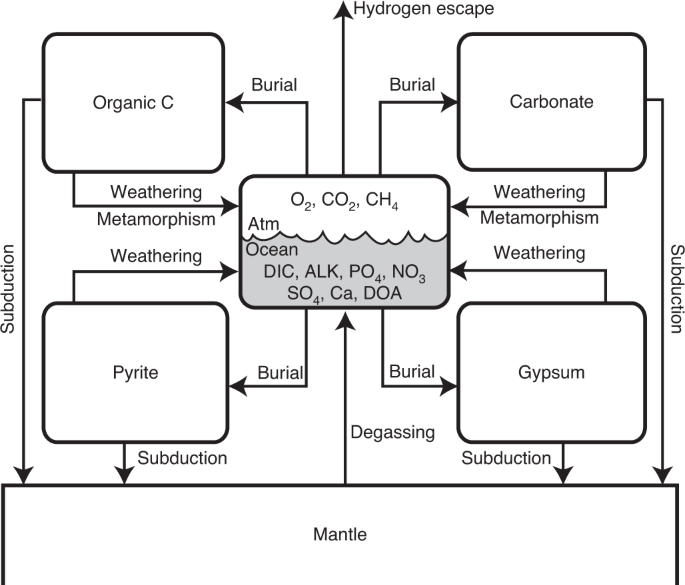
地球的现代大气层被高度氧化,是其表面生物圈的遥测信号。但是,地球大气中基于氧气的生物特征的寿命仍然不确定,尤其是在不久的将来。在这里,我们使用生物地球化学和气候相结合的模型来检查地球上富氧大气条件的可能时标。使用随机方法,我们发现,氧含量超过当前大气水平的1%以上的地球大气的平均未来寿命为1.08±1.44亿十亿年(1σ)。该模型预测,大气中的脱氧,大气中的O 2急剧下降至让人联想到古生地球的水平,很可能是在地球气候系统中出现潮湿的温室条件之前以及在地表水大量流失之前触发的。大气层。我们发现,未来的脱氧是太阳通量增加的必然结果,而其精确的时机却受到地幔与海洋-大气-地壳系统之间功率降低的交换通量的调节。我们的结果表明,碳酸盐-硅酸盐行星循环将趋于导致最终限制CO 2的生物圈和快速的大气脱氧,强调需要适用于弱氧化和缺氧的系外行星大气的健壮大气生物特征,并强调了大气有机霾的潜在重要性在行星可居住性的末期。
通过统计分析获得的数据可从https://doi.org/10.6084/m9.figshare.13487487.v1下载。本文提供了源数据。
4. Meadows,V。S.等。系外行星生物特征:了解氧气在其环境中的生物特征。天体生物学18,630–662(2018)。
5. Schwieterman,E. W.等。系外行星生物特征:对可远程检测到的生命特征的综述。天体生物学18,663-708(2018)。
6. Meadows,V.关于系外行星大气中O 2作为生物签名的思考。天体生物学17,1022–1052(2017)。
7.华兹华斯(R.&)。 Pierrehumbert,R.陆地宜居带行星上的非生物占主导地位的大气层。天体。 J.莱特785,L20(2014)。
8. Gao P.,Hu R.,Robinson T. D.,Li C.& Yung,Y. L.干燥的M矮系系外行星上CO 2气氛的稳定性。天体。 806,249(2015)。
9. Luger,R.& 巴恩斯河(Barnes,R.)在m个小矮星的宜居区域中,行星上极度失水和非生物O 2积累。 天文生物学15,119–143(2015)。 10.Prentice,I.C。等。 见《气候变化2001:科学基础》(eds Houghton,J. T.等人)183–238(IPCC,剑桥大学出版社,2001)。 11.里昂·T·W·莱恩哈德·C·T·安培。 新泽西州普拉纳夫斯基。地球早期海洋和大气中氧气的上升。 自然506,307–315(2014)。 12.Lenton,T.M。等人。 最早的陆地植物创造了现代水平的大气氧气。 程序。 纳特·阿卡德(Natl Acad)。 科学 美国113,9704–9709(2016)。 13.Krause,A.J。等人。 古生代大气的逐步充氧。 纳特 公社 9,4081(2018)。 15.T.M. Lenton,M.Crouch,M.Johnson,M.Pires,N。 多兰(L. Dolan)。第一批工厂冷却了奥陶纪。 纳特 Geosci。 5,86-89(2012)。 16. K. Caldeira,& Kasting,J. F.重新审视了生物圈的寿命。 Nature 360,721–723(1992)。
17. Lovelock,J。E.&惠特菲尔德,M。生物圈的寿命。自然296,561–563(1982)。
18. S.弗兰克(Franck,S.),布纳纳马(C. Bounama)& von Bloh,W.未来生物圈灭绝的原因和时机。生物地球科学3,85-92(2006)。
19. S. Franck,S。Kossacki,K。J.,W。Bloth,W。& Bounama,C。全球碳循环的长期演变:目前全球表面温度的历史最低值。 Tellus B 54,325–343(2002)。
20. O'Malley-James,J. T.,Greaves,J。S.,Raven,J。A.& Cockell,C. S. Swansong生物圈:生命的避难所和可居住寿命快结束时,地球行星上的新型微生物生物圈。诠释J. Astrobiol。 12,99-112(2013)。
21.沃克,J。C. G.,海斯,P。B.& Kasting,J. F.负反馈机制,用于长期稳定地球的表面温度。 J.地球物理。 Res。 86,9776–9782(1981)。
22. Kasting,J. F.失控和潮湿的温室气体以及地球和金星的演变。伊卡洛斯74,472–494(1988)。
23.Leconte,J.,Forget,F.,Charnay,B.,Wordsworth,R。珀蒂埃(A. Pottier)。增加了类似地球的行星上温室失控的日晒阈值。自然504,268–271(2013)。
24.克莱尔(M. W.),卡特琳(D. C.)和Zahnle,K. J.大气氧上升的生物地球化学模型。地球生物学4,239–269(2006)。
25.Reinhard,C.T。等。 cGENIE地球系统模型中的海洋和大气甲烷循环。 Geosci。模型开发讨论。 https://doi.org/10.5194/gmd-2020-32(2020)。
26. B.&B。 Goldblatt,C.高浓度混合温室气体的辐射强迫。地理学。 Res。来吧41,152–160(2014)。
27. Holland,H. D.火山气体,黑烟和巨大的氧化事件。 Geochim。宇宙猫Acta 66,3811–3826(2002)。
28. Arney,G。等人。浅橙色点:朦胧的太古宙地球的光谱和可居住性。天体生物学16,873–899(2016)。
29. Haqq-Misra,J. D.,Domagal-Goldman,S. D.,Kasting,P. J.& J. F. Kasting。一个为太古宙地球改建的朦胧甲烷温室。天体生物学8,1127–1137(2008)。
30. Trainer,M。G.等。早期地球大气中的烟雾弥漫:天上的甘露。天体生物学4,409–419(2004)。
31.Jenkins,J.M。等。开普勒452b的发现和验证:一颗G2星可居住区的1.6 R⊕超地球系外行星。阿斯特隆J.150,56(2015)。
32.Mullally,F.,Thompson,S.E.,Coughlin,J.L.,Burke,C.J。如果没有独立检测,就不应确认罗夫·J·F·开普勒的类地行星:开普勒452b案。阿斯特隆J.155,210(2018)。
33.麦肯齐(F. T.&)。 Kump,L. R.反向风化,粘土矿物形成和海洋元素循环。 Science 270,586–586(1995)。
34. Isson,T. T.& Planavsky,N. J.逆向气候作为海洋pH和行星气候的长期稳定剂。自然560,471–475(2018)。
35.沃尔夫,E.T。,希尔兹,A.L。,科帕帕拉普,R.K.,Haqq-Misra,J。 Toon,O. B. Toon的气候和可居住性的约束,这些约束来自于一般环流模型确定的类地球系外行星。天体。 J.837,107(2017)。
36. Gough,D。O.太阳内部结构和光度变化。索尔物理74,21-34(1981)。
37. S. A.的Wilde,J. W.的Valley,W. H.的Peck& A. Graham,C.来自碎石锆石的证据表明,在4.4 Gyr以前地球上存在大陆壳和海洋。自然409,175–178(2001)。
38. Reinhard,C.T.,Olson,S. L.,Schwieterman,E. W.& Lyons,T.W. W.远程寿命检测的虚假否定抗损伤:早期地球的课程。 Astrobiology 17,287-297(2017)。
39. Schwieterman,E.W.,Reinhard,C.T.,Olson,S.L.,Lyons,T.W. UV功能与下一代空间望远镜识别外产的能力的重要性。 [白皮书以回应国家科学院宇宙宇宙博士救生人员的反馈征求征求意见](2018年)。
40. Hayes,J. M.& Waldbauer,J.R。碳循环和相关的氧化还原过程通过时间。菲尔。跨。 R. SoC。 B 361,931-950(2006)。
41. Eguchi,J.,Heales,J.& Dasgupta,R.通过深循环和增强碳脱气的碳化和Lomagundi事件。 NAT。地质。 13,71-76(2020)。
42.埃文斯,K。A.俯冲区的氧化还原预算。地球科学。 Rev. 113,11-32(2012)。
43.卡杜达,S。,坎丁,D.C.C.,Nicklas,R.W.,Puchtel,I.& Anbar,A。D. D. Mantle数据意味着可氧化的火山气体的下降可能引发了巨大的渗透。 NAT。安排。 11,2774(2020)。
44.李,C.-t. A.等人。与大洲的生长有关的大气氧气两步升高。 NAT。地质。 9,417-424(2016)。
45. Ozaki,K.,Reinhard,C.T.&塔吉卡,E.一个缓慢的中间古代生物圈及其对地球氧化还原平衡的影响。地质学17,3-11(2019)。
46. Ozaki,K.,塔吉卡,E.,Hong,P. K.,Nakagawa,Y.& Reinhard,C.T.原始光合作用对地球早期气候系统的影响。 NAT。地质。 11,55-59(2018)。
48. Bergman,N.M.,Lenton,T. M.& Watson,A. J.Copse:在Phanerozoonooio的生物地球化学循环的新模型。是。 J. SCI。 304,397-437(2004)。
49. Laakso,T. A.& Schrag,D. P.限制限制。球。生物良果。 CYCLES 32,486-496(2018)。
50. Laakso,T. A.& Schrag,D. P. Preambrian氛围中的甲烷。地球星球。 SCI。吧。 522,48-54(2019)。
51.甘顿,迪恩,迪恩斯,S. J.&米尔斯,B. J.W.Copse重新加载:在Phanozoonooio的生物地球化学循环的改进模型。地球科学。 Rev. 178,1-28(2018)。
52.海因,M.& Sand-Jensen,K.Co 2增加了海洋主要生产。 Nature 388,526-527(1997)。
53. U. Riebesell,D。A.的Wolf-Gladrow。 Smetacek,V.海洋浮游植物生长速率的二氧化碳限制。 Nature 361,249–251(1993)。
54. Goldblatt,C.,Lenton,T. M.& Watson,A. J.大气中的氧气和大氧化的双稳性。自然443,683–686(2006)。
55. Galbraith,E. D.& Eggleston,S.在过去800,000年中对大气中CO 2浓度的下限。纳特Geosci。 10,295–298(2017)。
56. M. Pagani,K。Caldeira,R。& Berner。 Beerling,D. J.在过去的2400万年中,陆地植物在限制大气CO 2下降中的作用。自然460,85-88(2009)。
57.伯林(Berling),伯纳(Berner),麦肯齐(F.T.),哈特富特(M. B.)在过去的4亿年中,Pyle,J. A.甲烷与CH 4相关的温室效应。是。 J.科学309,97–113(2009)。
58. Canfield,D. E.地表硫储层的演化。是。 J.科学304,839–861(2004)。
59.Lécuyer,C.&里卡德(Y. Ricard),Y。铁的长期通量和预算:对地球地幔和大气的氧化还原态的影响。地球行星。科学来吧165,197–211(1999)。
60. Catling,D. C.& Kasting,J.F.《居住和无生命世界的大气演变》(剑桥大学出版社,2017年)。
61. Williams,D. M.& J. F. Kasting,高倾斜度的可居住行星。伊卡洛斯129,254–267(1997)。
62. Pierrehumbert,R. T.,Abbot,D. S.,Voigt,A.&科尔,D。新元古代的气候。安努牧师地球行星。科学39,417–460(2011)。
63. Goldblatt,C.,Robinson,T. D.,Zahnle,K. J.& Crisp,D。失控温室气候的模拟辐射下限低。纳特Geosci。 6,661–667(2013)。
64. Kopparapu,R. K.等。主序恒星周围的宜居带:新的估计。天体。 J.765,131(2013)。
65. Abe,Y.,Abe-Ouchi,A.,Sleep,N. H.& Zahnle,K. J.干燥行星的可居住区域限制。天体生物学11,443–460(2011)。
66. Krissansen-Totton,J.,Arney,G. N.& Catling,D. C.用地质碳循环模型限制早期地球的气候和海洋pH值。程序。纳特·阿卡德(Natl Acad)。科学美国115,4105-4110(2018)。
67. Krissansen-Totton,J.& Catling,D. C.使用反地质碳循环模型来限制气候敏感性以及大陆和海底风化。纳特公社8,15423(2017)。
68. Graham,R. J.& Pierrehumbert,R.大陆硅酸盐风化的热力学和高能极限极大地影响了潮湿多岩石世界的气候和可居住性。天体。 J.896,115(2020)。
69. Winnick,M. J.& Maher,K.硅酸盐风化的CO 2热力学极限与硅酸盐风化反馈强度之间的关系。地球行星。科学来吧485,111–120(2018)。
70. Royer,D. L.,Donnadieu,Y.,Park,J.,Kowalczyk,J.& Goddéris,Y.长期地球化学模型GEOCARBSULF对CO 2和O 2估计值的误差分析。是。 J.科学314,1259-1283,https://doi.org/10.2475/09.2014.01(2014)。
71.Ibarra,D.E。等。模拟陆地植物演变对硅酸盐风化的影响。是。 J.科学319,1-43(2019)。
72. Moulton,K. L.& Berner,R. A.量化植物对风化的影响:冰岛的研究。地质26,895–898(1998)。
我们感谢E. Tajika,Y。Sekine,S。Kadoya,Y。Watanabe,G。Arney,S。D. Domagal-Goldman和E. W. Schwieterman的有益讨论。 K.O.感谢由大学空间研究协会在与NASA接触下管理的NASA天文生物学计划中NASA博士后计划的支持。这项工作得到了JSPS KAKENHI资助号JP20K04066的支持。 C.T.R.感谢NASA天文生物学研究所的支持(批准号13-13NAI7_2-0027)。我们感谢NASA系外行星系统科学(NExSS)(授权号80NSSC19KO461)。
a,陆地初级生产的温度依赖性(V T)48。b,陆地初级生产的CO 2依赖性(V CO 2)。 c,对于在10 -2.5到10 -1.5(f UV)之间不同的c UV假定值,地面初级产品的UV辐照度依赖性(以大气O 2表示)。 d,海洋初级生产者的温度依赖性(f T ocn)24。灰色区域表示通过随机方法探索的这些因素的范围。
a,放气因子f G. b,侵蚀因子f R.对于生代而言,采用了原始模型48的时间演化,而未来的演化则通过改变振幅和周期来探索(方程(34)和(35))。灰色区域表示通过随机方法探索的那些因素的不确定范围。
假设先验分布均匀,但OLR和TOA的不确定性除外,其中不确定性假定为高斯概率密度函数(此处列出1σ)。
a,太阳光度(按现代值归一化)。 b,大气的化学成分。蓝色,绿色和橙色线分别表示O 2,CO 2和CH 4。 c,整体表面温度。 d,全球净初级生产(NPP)。实线表示参考曲线(n = 0.73,m = 1,G = a R = J s org = J s py = J s Fe-ox = J s carb = J d CO2 = J d H2S = J d红色=ΔOLR=ΔTOA= 0,G * = 1250 Emol,C * = 5000 Emol,PYR * = 200 Emol,GYP * = 200 Emol,c UV = 0.01,T ref = 1400 K,T ref land = 25 C, LIFE = 0.15,FERT = 0.4,ACT = 0.09,P min = 10 ppmv,RUN sil = 0.038,(C org / P org)缺氧= 20×(C org / P org)有氧),而虚线(f G = 1,f R = 1),点线(不带陆地植物),点划线((C org / P org)无氧= 2×(C org / P org)有氧)和灰色(恒定太阳光度)线代表灵敏度实验(参数是在参考情况下设置的,否则注明)。潘= han生代。源数据
与扩展数据图4相同,而虚线(ACT = 0.135)和虚线(ACT = 0.05)表示灵敏度实验(参数在参考情况下设置,否则另有说明)。由于耐候性对大气CO 2水平,全球气候和生物圈响应的影响之间存在折衷关系,因此,未来寿命对陆地可湿性的不确定性不敏感。潘= han生代。源数据
a,太阳光度(按现代值归一化)。 b,大气的化学成分。蓝色,绿色和橙色线分别表示O 2,CO 2和CH 4。 c,整体表面温度。 d,全球净初级生产(NPP)。潘= han生代。
a,海洋中的P浓度,(b)海洋中的SO 4 2-含量,(c)地壳有机碳,(d)地壳碳酸盐碳,(e)地壳黄铁矿硫,(f)地壳石膏硫。
a,控制海洋生物圈活动的温度因子,T ref,(b)控制陆地生物圈活动的温度因子,T ref土地,(c)陆地植物的最低CO 2水平,P min,(d)径流因子 ,RUN sil,(e)活化能因子,ACT,(f)埋藏的缺氧沉积物的C org / P org比,(C org / P org)缺氧,(g)陆地植物的CO 2施肥因子,FERT, (h)无植被区的风化因子LIFE,(i)陆地生物圈的UV因子,c UV。 未来寿命对这些不确定的参数不敏感(参见图3b)。